
Sea Squirt
Tadpole larva, siphon-powered adult
Tadpole larva, siphon-powered adult

Sting-powered drifters of the sea

Pale eyes. Loud calls. City-smart flocks.

Cold-water royalty of the seafloor

More than night flyers
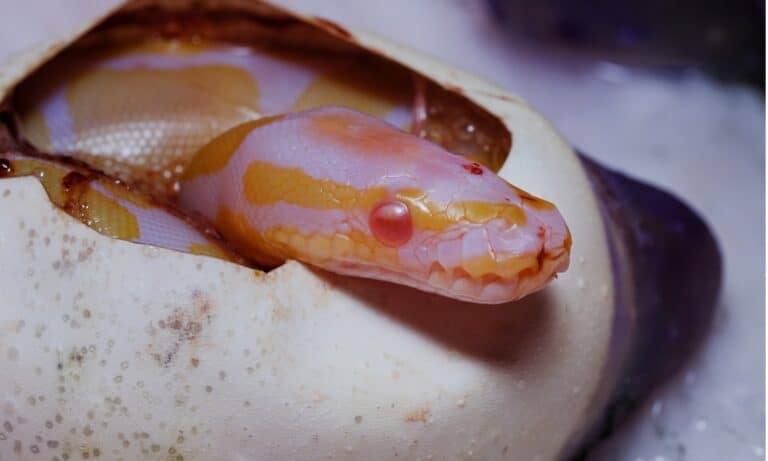
Soft lavender, royal serpent

Stinging "flowers" of the seafloor

Bright colors, borrowed chemistry

Coins of the seafloor, built to burrow

Sprint hunters of sunlit ground
In animal coloration, purple refers to hues with dominant spectral components at short wavelengths (blue-violet) combined with longer-wavelength red components, producing an appearance between blue and violet/magenta to human vision. In many animals, "purple" is produced not by a single purple pigment but by structural coloration and/or mixtures of pigments (often including porphyrins) and optical layering in feathers, scales, or skin.
Purple is one of the most striking-yet comparatively rare-colors in the animal kingdom. To human observers it spans a range from soft lavender and lilac to deep royal purple and violet, often with an iridescent sheen that shifts as the animal moves. Because true spectral "purple" does not correspond to a single wavelength, animals frequently achieve purple through clever physical and biochemical solutions: microscopic surface structures that scatter and interfere with light, or pigment combinations that shape reflected wavelengths into a purple impression.
Where it occurs, purple can serve multiple ecological roles. In birds and many insects, iridescent purples are commonly linked to structural coloration, producing flashes used in courtship displays, territorial signaling, or species recognition. In other cases, purple hues may contribute to camouflage in flower-rich or twilight environments, or function as warning or startle coloration when revealed suddenly.
Purple's scarcity in many vertebrate groups-especially mammals-reflects both biological constraints (limited pigment types and lower prevalence of certain light-manipulating structures) and sensory ecology (what conspecifics can detect and how signals transmit through habitat lighting). As a result, purple animals often stand out as examples of specialized evolution, where physics (nanostructures and optical layering) and chemistry (unusual pigments such as porphyrins) converge to create an uncommon visual effect.
On animals, "purple" typically reads as a blue-red/violet hue that can look lavender, lilac, magenta-leaning violet, or deep royal purple depending on lighting and the animal's surface microstructure. True, pigment-based purple is relatively rare in many vertebrates; purple appearances often come from: - Structural coloration (nano/microstructures in feathers, scales, cuticle) that selectively reflect short wavelengths and can shift with angle (iridescence), producing violet-to-purple flashes. - Pigments such as porphyrins (notably in some birds) that can appear reddish-violet under daylight and may fluoresce under UV. - Layering/optical mixing: a red/pink pigment over a blue-structural base (or vice versa) can yield a purple impression. In feathers, purple often appears as a glossy, metallic sheen or as velvety patches with high saturation; in scales or insect cuticle it can be mirror-like and strongly angle-dependent. In skin (less commonly), it may appear as a muted mauve or bruised-plum tone, sometimes influenced by blood perfusion plus scattering. In many species, purple areas are most vivid in direct sun and can dull to grayish-violet in shade; iridescent purples can "blink" on/off with movement.
Not a single spectral wavelength for most viewers: "purple" is largely a non-spectral hue (a percept formed by combining long- and short-wavelength stimulation). Violet (spectral edge that can contribute to purple impressions) is roughly ~380-450 nm; many animal "purple" appearances involve reflectance peaks in the blue/violet plus longer-wavelength (red) components or broad reflectance with structural enhancements.
Approx. #C8A2C8 (lavender) through #800080 (purple) to #4B0082 (indigo) and #3F00FF (electric/blue-violet); deeper "royal" purples often fall around #5B2C83-#6A0DAD.
Humans: Humans are trichromats and typically perceive "purple" as a mix of red and blue signals (a non-spectral percept). Many animal purples-especially iridescent ones-change dramatically with viewing angle and lighting, so humans may see them shift from violet to blue or even appear black when not catching light. Birds: Most birds are tetrachromats and often have ultraviolet-sensitive or violet-sensitive cones. Many "purple" patches that look modest to humans can be much more saturated and higher-contrast to birds because UV reflectance adds an extra channel; conversely, some purples that rely on human red+blue mixing may map differently in avian color space and be perceived as a distinct hue not equivalent to human purple. Insects (e.g., bees, butterflies): Many insects are UV-blue-green trichromats and have little or no sensitivity to long-wavelength red. Human 'purple' that depends on red components may not read as purple at all; it may appear as blue/violet or as a different category entirely. Structural purples with strong UV/blue reflectance can be highly salient. Reptiles/amphibians/fish: Visual systems vary widely (often 3-4+ cone types; some extend into UV). Purple signals may be enhanced or shifted depending on UV sensitivity and the water/light environment for aquatic species. In water, long wavelengths attenuate quickly, so purple with a strong red component may look bluer at depth. Mammals: Many mammals are dichromats (limited red-green discrimination). Purple often collapses toward blue or gray because the 'red' component contributing to human purple may not be distinctly encoded. As a result, purple markings may be less communicative to many mammals than to birds or UV-capable species.
Low-to-moderate saturation, high lightness; often seen as a soft wash on feathers or skin where pigment is sparse or mixed with white structural scattering. Can look grayish in shade and more rosy in warm light.
Purple shifted toward magenta; can arise from red pigments combined with bluish structural reflection. Often reads as more "floral" and can be especially conspicuous to species with good red sensitivity.
High saturation with a strong short-wavelength component; commonly produced by structural coloration in feathers/scales/cuticle. Frequently appears brightest in direct light and may shift toward blue as viewing angle changes.
Very dark, rich purple with low lightness; may be pigment-heavy or a darker structural effect. In dim light it can collapse toward near-black with only a faint violet cast.
Angle-dependent purple highlights over a darker base (black/brown). Seen in many iridescent feathers and insect exoskeletons; the hue can shift purple→blue→green depending on geometry and illumination.
Surfaces with strong ultraviolet reflectance plus visible blue/violet can appear merely bluish or grayish to humans but stand out as highly chromatic to UV-sensitive animals (e.g., many birds, insects).
Pigment-driven purple that can look wine/mulberry under daylight; may show fluorescence under UV. Often less angle-dependent than structural purple but can fade with light exposure.
Not a chemical pigment but nanoscale architecture (e.g., quasi-ordered spongy keratin in feather barbs, multilayer lamellae in butterfly scales, cuticular photonic crystals) that preferentially reflects violet/blue light. When paired with red pigments or dark backing, the perceived color can become purple/violet.
Black/brown melanin that does not create purple by itself, but commonly underlies structural reflectors to absorb stray light, increasing saturation and shifting appearance toward deeper violet/purple. In some iridescent tissues, melanosomes also participate as layers in interference stacks.
Diet-derived red/orange pigments. When combined with structural blue/violet reflection (or overlaid on it), they can yield a composite purple/magenta tone (additive/subtractive mixing depending on tissue optics).
Tetrapyrrole pigments that can appear reddish, brownish, or purplish depending on concentration, binding environment, and lighting; often show strong fluorescence under UV. In some birds, porphyrins contribute to reddish to pinkish/purplish hues in feathers and can combine with structure/melanin to look violet.
In insects and some vertebrates (notably many reptiles/amphibians/fishes via chromatophores), pteridines provide red/orange components. Together with structural blue (iridophores) they can yield purple/violet effects in skin or scales.
Many "purple" appearances are emergent: violet structural reflection + red pigment overlay, or angle-dependent iridescence that shifts toward purple under certain illumination backgrounds. No single pigment is responsible.
Purple coloration is most adaptive as a high-information visual signal (mate attraction, social communication, and species recognition), especially when produced structurally and displayed in good light to color-discriminating receivers. Secondary benefits can include predator deterrence (warning/mimicry) or niche-specific camouflage (flower/algae/low-light darkening). Costs-conspicuousness to predators, production/maintenance costs, and variable appearance under different lighting/angles-tend to restrict purple to taxa/environments where signaling benefits outweigh detection risk.
Purple can act as a high-salience visual signal because it is uncommon in many natural backgrounds and can appear vivid when produced by structural coloration (iridescence) or by pigments (e.g., porphyrins). It may convey state (health, diet, parasite load) if the color is costly to produce/maintain or degrades with wear.
Effectiveness: Most effective in well-lit, open or canopy-filtered environments where short-wavelength light is available and receivers have UV/blue sensitivity (many birds, some reptiles, insects). Less effective at dusk/night, in turbid water, or deep forest shade where purple shifts toward dark/black; also less effective if the intended viewers lack relevant cone sensitivity.
Purple ornamentation (patches, crests, dewlaps, facial skin, plumage gloss) can function as a mate-attracting trait or in male-male competition. Structural purple often correlates with microstructure quality, making it a potentially honest indicator.
Effectiveness: Highly effective when mate choice relies on color discrimination and display occurs in direct sun or bright diffuse light; effectiveness increases with motion/display behaviors that accentuate iridescence. Reduced effectiveness where predation pressure penalizes conspicuous colors, where displays occur in low light, or where habitat colors include abundant purple flowers/fruits that reduce contrast.
Distinctive purple marks can help conspecifics rapidly identify species or morphs, reducing hybridization and misdirected aggression, especially in communities with many similarly colored taxa (e.g., bright passerines, reef fish, butterflies).
Effectiveness: Effective in biodiverse habitats with many look-alikes and in social species needing quick recognition. Less effective in low visibility (fog, murky water) or when purple is only visible at certain angles (strong iridescence) unless paired with pattern/shape cues.
When combined with high contrast (black/yellow/white) or glossy iridescence, purple can contribute to aposematic signaling, indicating toxicity, unpalatability, spines, or defensive chemicals. Its rarity may enhance novelty-based avoidance in predators.
Effectiveness: Most effective for visually hunting predators with color vision and learning capacity (birds, some lizards, fish). Less effective against predators relying on olfaction/tactile cues, in dim habitats, or when purple appears dark and loses conspicuousness.
Purple can be used by harmless species to mimic defended species that include violet/iridescent elements, or by predators/parasites to resemble attractive resources (flowers/fruit) and lure prey (aggressive mimicry).
Effectiveness: Effective where the model is common and predators have learned the association; also effective in pollination/flower-mimic contexts for insects. Weaker where models are rare, where purple varies strongly with viewing angle (making mimicry inconsistent), or where predators generalize poorly across hue/brightness.
Although uncommon as a pure background color, purple can provide camouflage via (1) background matching in habitats with violet flowers, purple algae/coralline surfaces, or berry-rich vegetation; (2) countershading/iridescence that breaks up outlines; (3) appearing dark (nearly black) in low light, aiding concealment.
Effectiveness: Effective in flower- and berry-rich landscapes, reef/algal environments, and dusk/understory conditions where purple reads as dark. Generally less effective in green-dominated foliage, sandy deserts, or clear open water where purple creates strong contrast.
Dark violet/purple surfaces may absorb more solar radiation than pale colors, potentially aiding warming in cool environments; conversely, iridescent structural purple can reflect portions of near-IR/visible depending on microstructure, sometimes moderating heat gain.
Effectiveness: Useful for basking ectotherms in cool, sunny conditions (dark purple warms faster). Potentially disadvantageous in hot, exposed habitats due to overheating unless behavior (shade use, posture) compensates. Iridescent purples with high reflectance can be neutral or mildly beneficial in variable climates.
Purple coloration can be linked to chemical/physical protection: porphyrin-based pigments and related compounds can be associated with antimicrobial properties in some taxa, while structural colors may coincide with feather/scale microstructures that increase water repellency or reduce wear. Purple-black can also reduce UV damage to underlying tissues by increasing absorption.
Effectiveness: Context-dependent and often indirect: more effective where UV exposure is high or microbial load is elevated (humid habitats) and when pigment/structure confers measurable tissue benefits. Less effective if pigments photodegrade quickly (some porphyrins) or if protection relies mainly on behavior rather than color/structure.
Purple is commonly sexually dimorphic when used in sexual selection: males often exhibit more saturated, larger, or more iridescent purple patches (plumage, throat/crest, dewlaps, facial skin), while females are duller, browner/gray, or show reduced patch size to lower predation risk during nesting/gestation. In species where purple functions mainly in species recognition or warning, both sexes may be similarly colored (monomorphic), with dimorphism expressed more in intensity/iridescence than in pattern presence/absence.
Purple coloration can affect conservation in both helpful and risky ways. Its rarity and visual appeal can increase public interest, donations, and outreach effectiveness (a "flagship effect"), especially for charismatic taxa like birds, butterflies, and reef fish. However, conspicuous coloration may also increase detectability to predators and humans, potentially raising poaching/collection pressure when purple morphs are targeted for the pet/ornamental trade. In some species, purple hues arise from structural coloration sensitive to habitat quality (e.g., feather microstructure linked to condition, diet, or parasite load), so changes in purple intensity can serve as a bioindicator for ecosystem health-useful in monitoring programs. Conversely, when purple appearance is caused by contaminants, disease, or abnormal pigmentation pathways (e.g., porphyrin-related coloration in some taxa), sightings may signal environmental stress, requiring careful communication to avoid misinformation while prompting investigation. Conservation planning may therefore need: (1) trade regulation and enforcement for high-value morphs, (2) guidance for citizen scientists to document without disturbing/collecting, and (3) research to determine whether purple expression correlates with fitness or habitat degradation.
Many "purple" animals aren't using purple pigment at all-the color can be an optical trick created by microscopic structures that reflect violet/blue while absorbing other wavelengths.
A lot of purple you see in birds and butterflies is angle-dependent: the same animal can look deep violet from one view and shift toward blue or even dark/black from another.
Some organisms can appear purple simply because they lack other pigments: removing or reducing yellow/red pigments can make underlying blues look violet-ish to our eyes.
Purple can be a warning color in the sea even when it looks like "royal" coloration to us-at depth, reds disappear first, and purples may read as dark or high-contrast rather than bright.
Certain "purple stains" in nature come from chemistry outside the animal (diet, symbiotic microbes, or surface films) rather than the animal producing a purple color itself.
If bright red is the "loud" end of warm colors on land, vivid purple is like a rarer, more specialized "loud signal"-common in insects and marine life, much less so in birds, and almost never in mammals.
Structural purple is like a tiny natural prism on the animal's surface: instead of dyeing the body purple, the body is built like a light-manipulating surface that *creates* purple from reflected light.
In underwater habitats, a purple animal can function like a "high-contrast silhouette" rather than a bright neon sign-because water filters light and changes which colors stay visible.
Compared with blues, purples often require either more complex nanostructures (to shift reflected color toward violet) or additional absorbing pigments to "push" the hue away from pure blue.
Among vertebrates, purple is one of the rarest "true" body colors-especially in mammals, where vivid purple is essentially absent in natural fur and skin.
Some marine invertebrates can display the most saturated "electric purple" hues in nature thanks to structural coloration (nanostructures that scatter light), often appearing brighter than many pigment-based purples.
Purple sea urchins (e.g., *Strongylocentrotus purpuratus*) are among the most famously purple animals by sheer visibility and abundance along certain coastlines-an iconic "signature purple" species.
Sting-powered drifters of the sea
Cold-water royalty of the seafloor
More than night flyers

Hydraulic feet, star-shaped predators

Spines, jaws, and ocean power

Naked gills, wild skills.

Hover. Sip. Pollinate. Repeat.

Nature's greatest courtship show

Porous powerhouses of the sea
Tadpole larva, siphon-powered adult

Purple flash in the treetops

Savanna's living rainbow finch

A colony that sails and stings
Stinging "flowers" of the seafloor
Sprint hunters of sunlit ground

Tentacled tunnel-hunters of the tropics

Beaks that build beaches

Calico traveler of the North Pacific

Deep reef royalty in purple and gold

Small fish, big reef fireworks

Small crab, big attitude.

Nature's living metalwork
Soft lavender, royal serpent
Coins of the seafloor, built to burrow
Thank you for reading! Have some feedback for us?
We appreciate your help in improving our content.
Our editorial team will review your suggestions and make any necessary updates.
There was an error submitting your feedback. Please try again.